Axe 2. Structure et dynamique de membranes cellulaires et de nanoparticules lipidiques Responsables : Frédéric PRZYBILLA et Toshihide KOBAYASHI
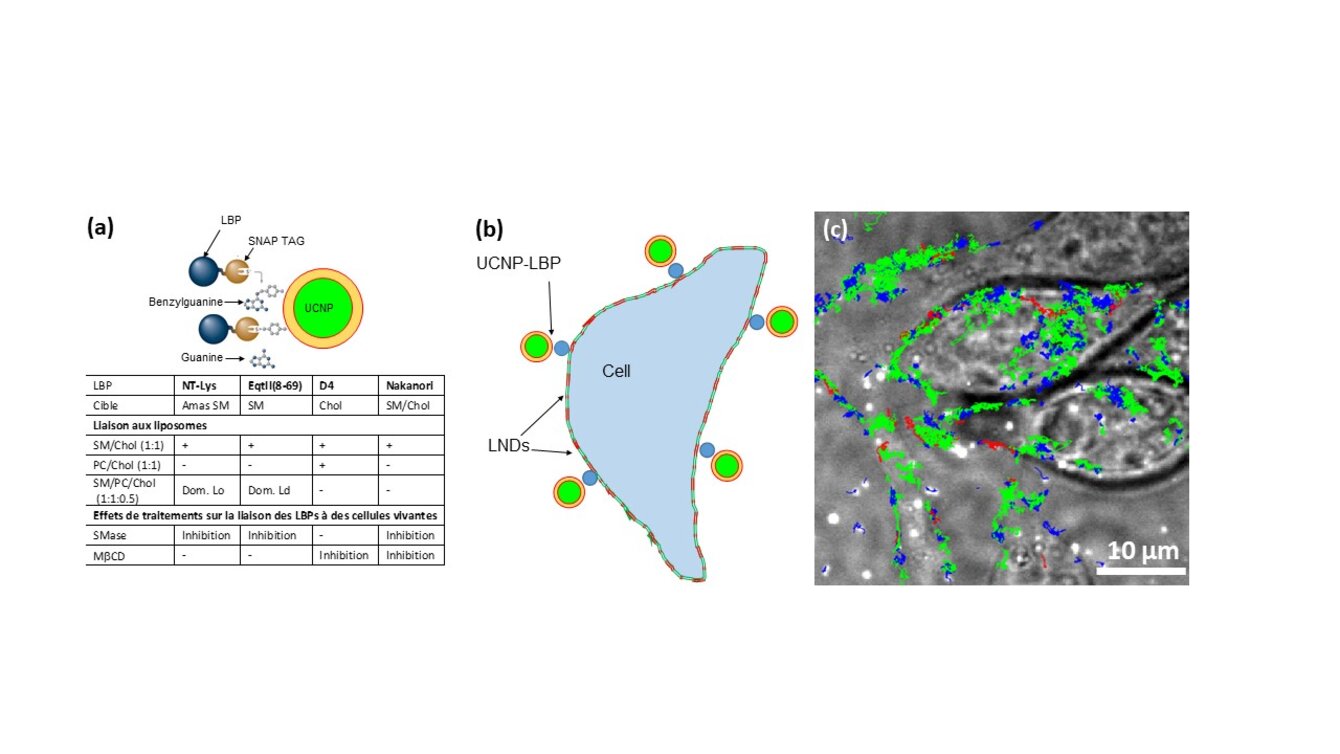
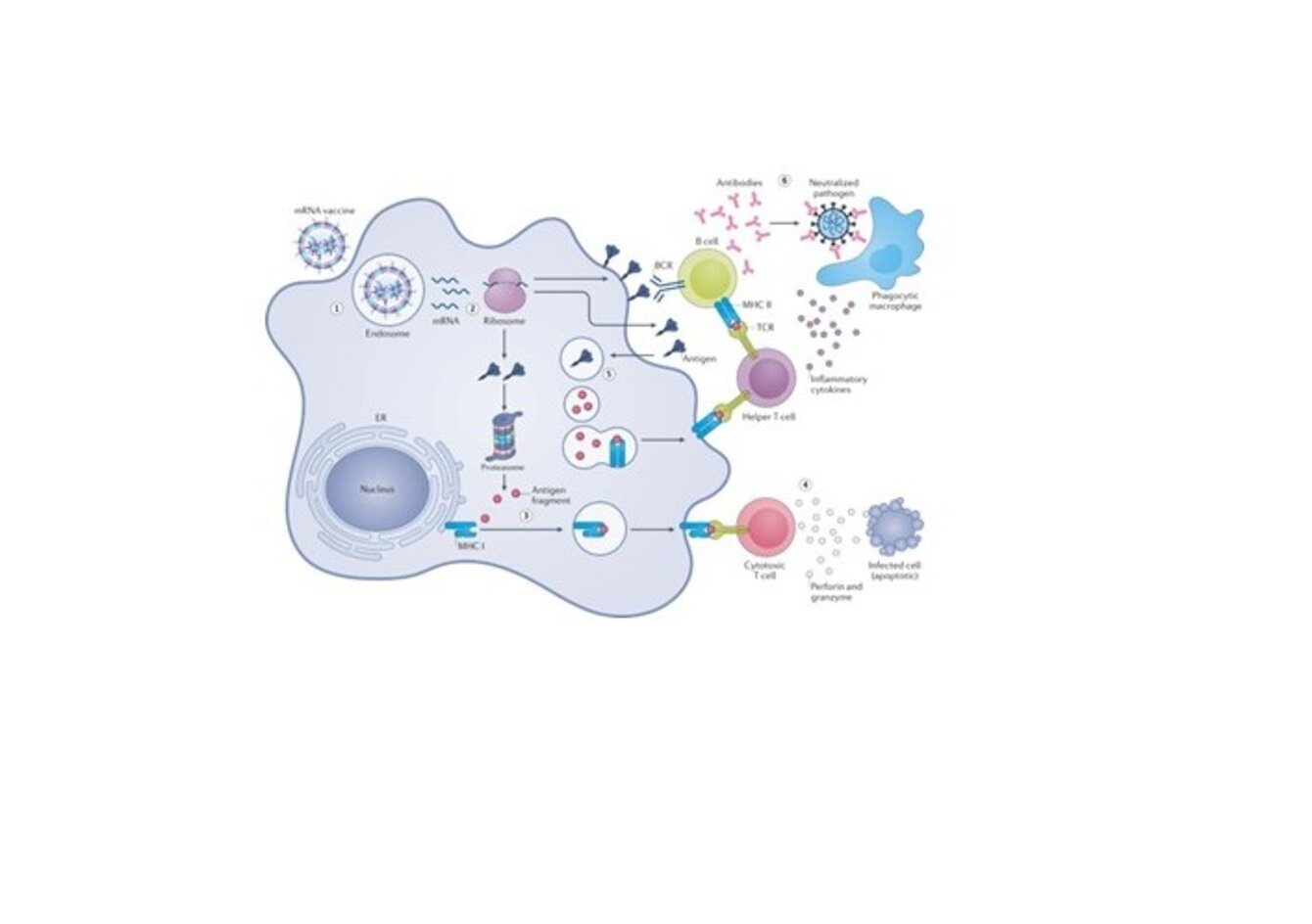
Au sein d’une membrane, la composition lipidique des feuillets externe et interne de la bicouche lipidique est différente et présente des hétérogénéités. La façon dont cette asymétrie/hétérogénéité est construite, maintenue et régulée reste mal comprise, tout comme son rôle. Nous avons développé une large palette d’outils innovants (sondes, protocoles, équipements) qui permet de caractériser et suivre ces organisations lipidiques in situ. En particulier, nous avons développé une série de sondes protéiques se liant spécifiquement à des lipides (LBPs pour lipid binding proteins) pour visualiser la distribution des lipides endogènes à l'échelle nanométrique.1,2 Notre objectif est d’utiliser ces LBPs pour étudier les mécanismes moléculaires régulant la distribution des lipides ainsi que leurs dynamiques. Nous couplerons également les LBPs à des nanoparticules à conversion ascendante (UCNPs pour up-converting nanoparticles) pour suivre la dynamique à long terme de nano domaines lipidiques (NDLs). Enfin, nous caractériserons la structure et le devenir intracellulaire de nanoparticules lipidiques encapsulant des ARNm (LNPs pour lipid nano-particles) utilisées en stratégie vaccinale.
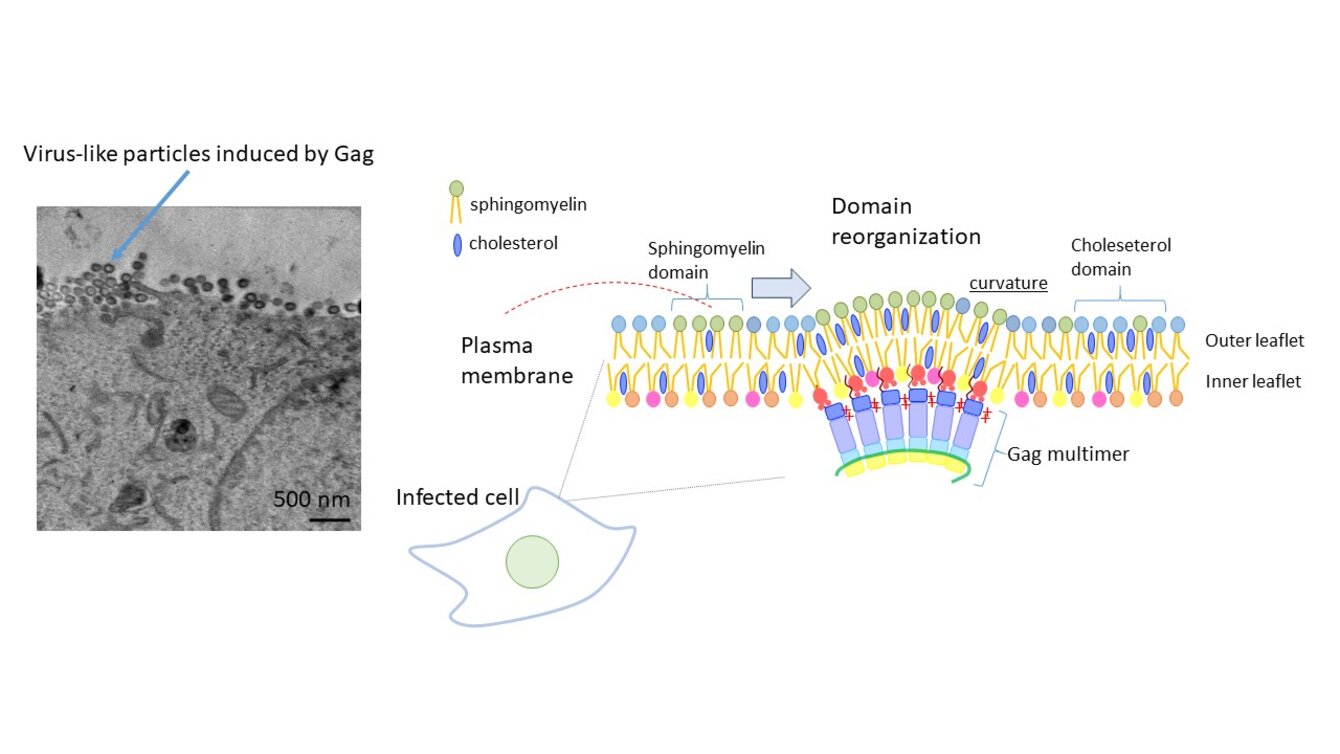
Découvrir comment VIH-1 assemble l’enveloppe lipidique qui lui permet d’entrer dans ses cellules cibles
L'enveloppe lipidique du virus de l'immunodéficience humaine de type 1 (VIH-1) est formée lors de l’étape de bourgeonnement au niveau de la membrane plasmique des cellules hôtes infectées. Il est intéressant de noter que le VIH-1 sélectionne un ensemble spécifique de lipides tels que la sphingomyéline (SM) et le cholestérol (Chol), qui sont importants pour l'activité du virus. La protéine virale Gag joue un rôle central dans le bourgeonnement du virus. Différentes techniques de microscopie quantitative et à super-résolution en combinaison avec des LBPs spécifiques de SM et de Chol ont révélé que lors de son expression dans une cellule, Gag recrute SM et Chol et induit la fusion de domaines riches en SM et en Chol.3 Le SM est distribué dans le feuillet externe de la membrane plasmique tandis que Gag se lie au lipide du phosphatidylinositol 4,5-bisphosphate (PI(4,5)P2) dans le feuillet interne. Ainsi, la communication entre les lipides du feuillet externe et interne est cruciale pour le recrutement des lipides par Gag. Cependant, on sait peu de choses sur la communication lipidique interbicouche dans les biomembranes. En utilisant la génétique moléculaire et diverses LBPs, nous étudierons les mécanismes moléculaires qui régulent la communication lipidique inter-feuillets. Notre étude découvrira comment le VIH-1 assemble son enveloppe lipidique et fournira également des informations essentielles sur la transduction du signal par la membrane.