Axe 3. Développements technologiques et méthodologiques Responsables : Frédéric PRZYBILLA et Ludovic RICHERT
Cet axe transversal vise à répondre aux besoins en microscopie du laboratoire et s’inscrit dans notre participation en tant qu’équipe R&D du noeud Alsace de France Bioimaging. Notre objectif est de développer de nouvelles modalités d’imagerie optique à la pointe de la technologie et de nouvelles sondes fluorescentes capables d’apporter des réponses décisives aux questions en biologie de l’équipe et de l’UMR.
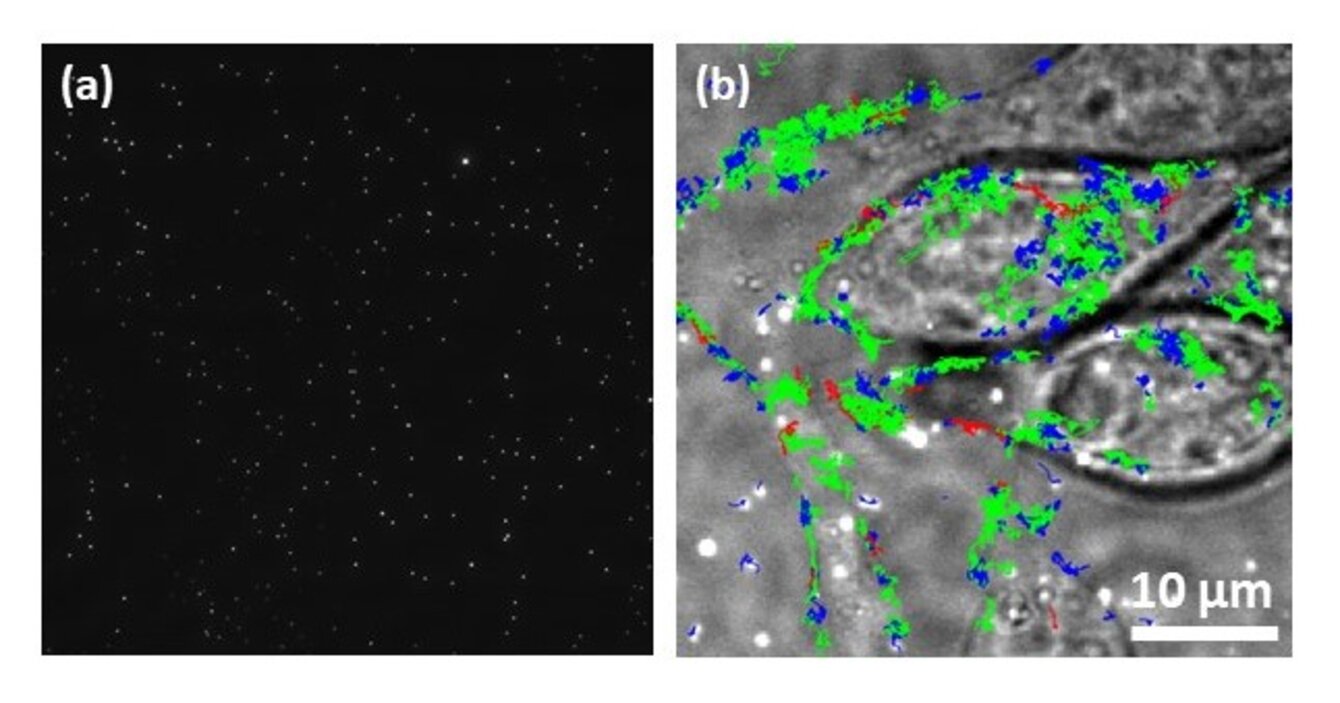
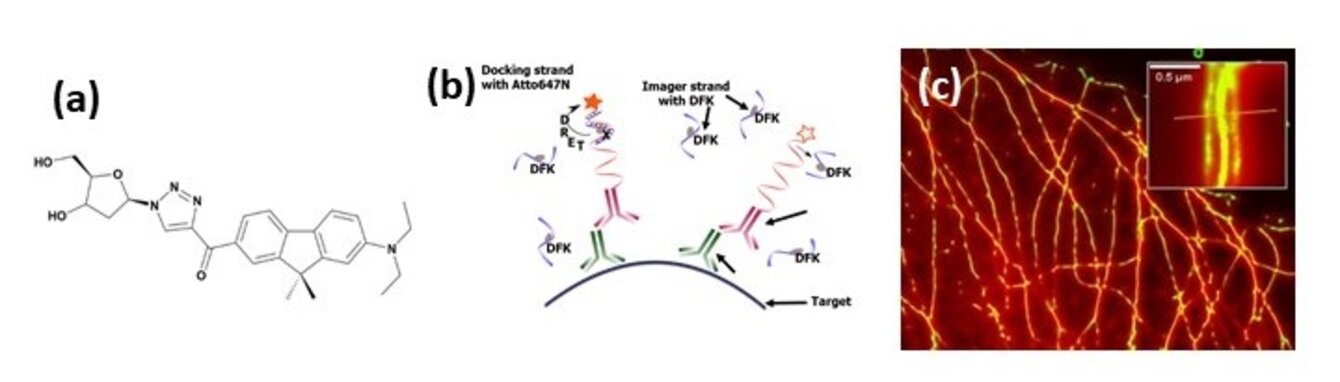
Nous avons acquis une forte expertise dans les approches quantitatives de microscopie : les F-Techniques (FLIM/FC(C)S)1 et les caractérisations à l’échelle de la molécule unique (smFRET (single molecule FRET), SPT (single particle tracking), SMLM (Single Molecule Localization Microscopy)2 pour l’étude des dynamiques et interactions biomoléculaires (protéine-protéine, acide nucléique/protéine, lipide/protéine). Après les phases de prototypage et validation des nouveaux instruments, ces derniers sont progressivement intégrés à la plateforme QuESt-IBiSA pour les rendre accessibles à la communauté scientifique académique et industrielle. Nous avons ainsi déjà développé un microscope multiphoton FLIM (fluorescence lifetime microscopy)/FCCS (fluorescence cross correlation spectroscopy),3,4 2 microscopes de super-résolution (3D PALM/dSTORM et un système de λ-PAINT),5,6 un microscope champ large et un microscope confocal dédié à la conversion ascendante de photons respectivement optimisé pour le SPT cellulaire et imagerie PLIM (phosphorescence lifetime imaging).7,8
Sur la base de cette expertise et des nouveaux besoins des autres axes de l’équipe et du laboratoire, plusieurs développements seront menés tant en optimisant les instruments existants qu’en développant de nouvelles approches.