Axis 2. Structure and Dynamics of Cell Membranes and Lipid Nanoparticles Leaders: Frédéric PRZYBILLA and Toshihide KOBAYASHI
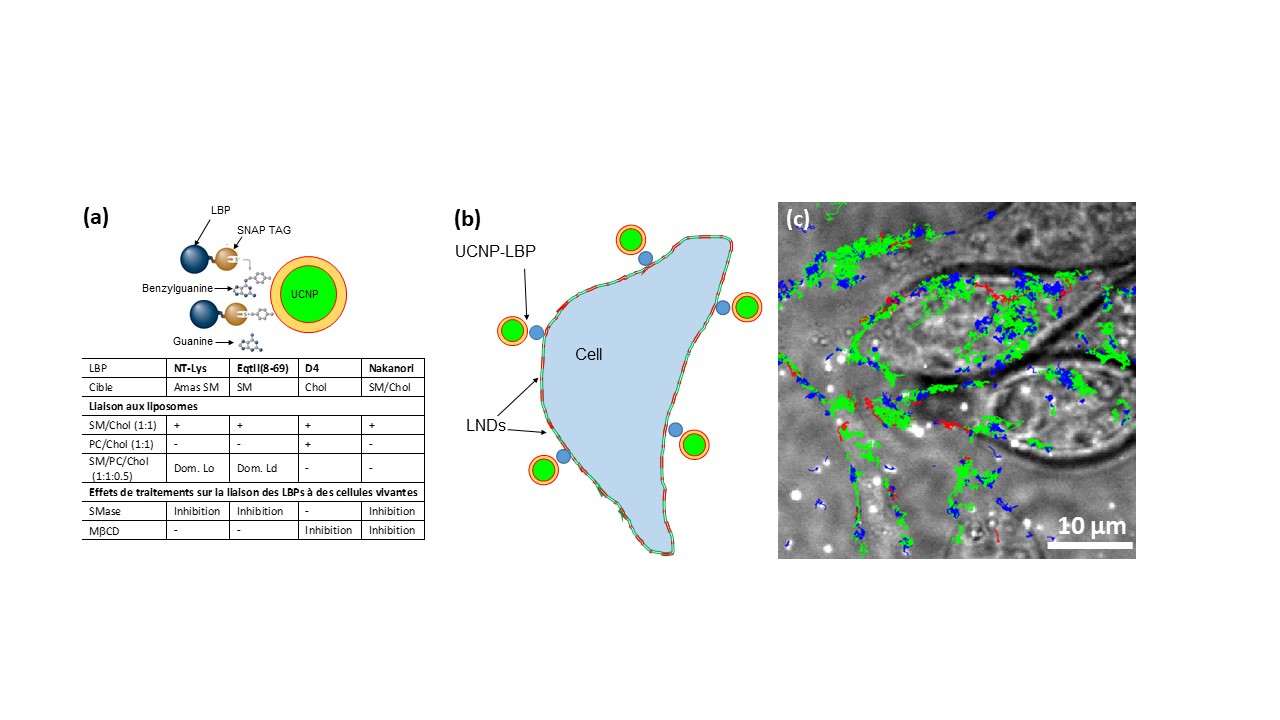
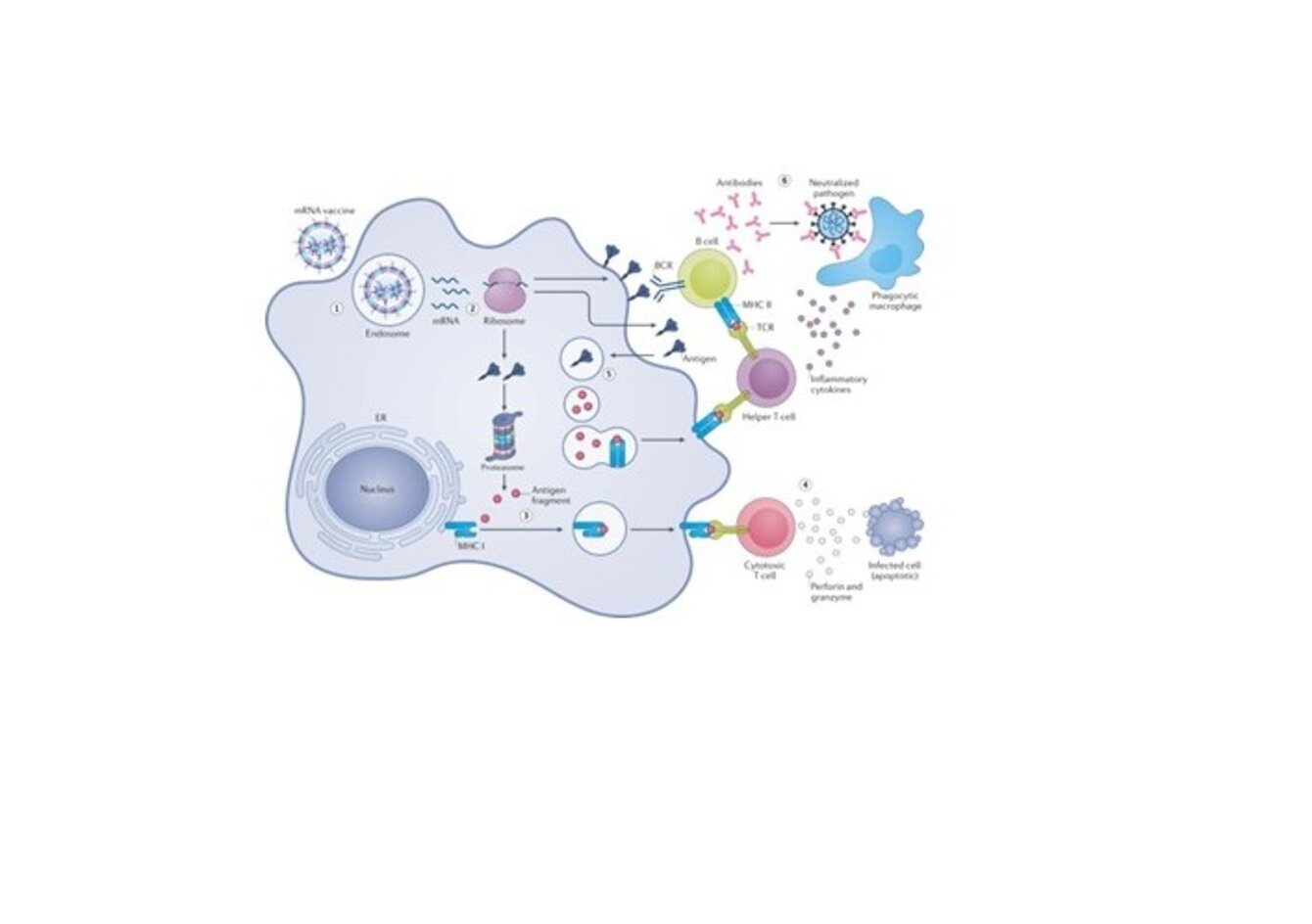
Our body contains thousands of different lipid species. These lipids are not randomly distributed in different organs and different organelles. Even in one membrane, the lipid composition of the outer and inner leaflets of the lipid bilayer is different. In addition, lipid distribution is heterogeneous in each leaflet. However, how this lipid organization is built, maintained and regulated remains poorly understood as is the physiological significance of lipid organization. We have been developing a wide range of innovative tools (probes, protocols, equipment) that allows the characterization of these lipid organizations in situ and at nanometer scale. In particular, a series of lipid binding proteins (LBPs) were designed and optimized to visualize the distribution of endogenous lipids at the nanoscale.1,2 Our objective is to use these LBPs to study the molecular mechanisms regulating the distribution of lipids as well as their dynamics. To follow lipid dynamics, we will couple our LBPs with the upconversion nanoparticles (UCNPs) approaches recently developed in the team to monitor the long-term dynamics of lipid nanodomains (NDLs). In addition, we will characterize the structure and intracellular fate of lipid nanoparticles encapsulating mRNAs (LNPs for lipid nano-particles) used in vaccine strategies.
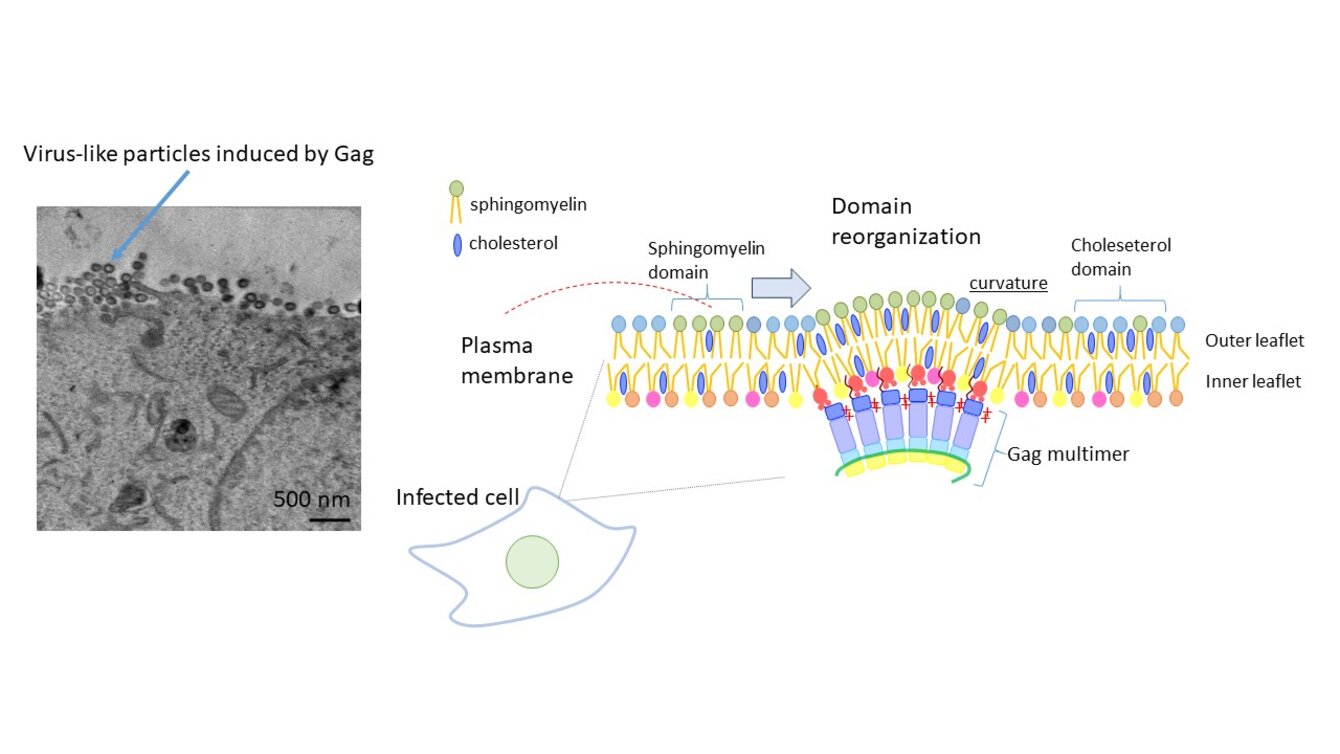
Uncovering how HIV-1 assembles its lipid coat, which allows the virus to enter cells
Human immunodeficiency virus type 1 (HIV-1) lipid envelope is obtained during budding from the plasma membrane of infected host cells. Interestingly, HIV-1 selects specific set of lipids such as sphingomyelin (SM) and cholesterol (Chol) that are important for virus activity. Viral Gag protein plays a central role in virus budding. Different quantitative and super-resolution microscopy techniques in combination with SM-specific and Chol-specific LBPs revealed that the expressed Gag recruits SM and Chol and induces fusion of SM-rich and Chol-rich domains.3 SM is distributed in the outer leaflet of the plasma membrane whereas Gag binds to the inner leaflet lipid phosphatidylinositol 4,5-bisphosphate (PI(4,5)P2). Thus, communication between outer and inner leaflet lipids is crucial for lipid recruitment by Gag. However, little is known about the interbilayer lipid communication in the biomembranes. Using molecular genetics and various LBPs, we will study molecular mechanisms that regulate interbilayer lipid communication. Our study will uncover how HIV-1 assembles its lipid coat. Our study will also provide essential information on membrane-mediated signal transduction.